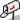|
Jean Benoist
“Les groupes sanguins des Canadiens français :
I. La région du Saguenay-Lac Saint-Jean”.
Un article publié dans la revue L'Anthropologie (Paris). Tome 73, no 1-2, 1969, pp. 49-76.
- INTRODUCTION
-
- I. LA RÉGION DU SAGUENAY-LAC SAINT-JEAN ET SON PEUPLEMENT
-
- II. LES GROUPES SANGUINS ABO ET RH
-
- A. L'échantillon.
- B. Les résultats.
-
- 1˚ Le système ABO.
- 2˚ Le système Rh.
- Comparaisons.
-
- III. LA RÉPARTITION RÉGIONALE DES GROUPES SANGUINS AU SAGUENAY-LAC SAINT-JEAN
-
- A. La division du territoire.
- B. Les groupes sanguins.
-
- 1˚ Système ABO.
- 2˚ Le système Rh.
- 3˚ Comparaison entre les régions.
-
- CONCLUSION
-
- BIBLIOGRAPHIE
-
-
- Appendice. Démographie des sous-populations du Saguenay-Lac Saint-Jean.
-
- Fig. 1. Situation géographique du Saguenay-Lac Saint-Jean
- Fig. 2. Les grandes étapes du peuplement
- Fig. 3. Répartition de 5 populations suivant la méthode de Streng : groupes sanguins ABO (explications dans le texte).
- Fig. 4. Les 15 subdivisions du Saguenay-Lac Saint-Jean
- Fig. 5. Groupes ABO : répartition des écarts significatifs dans la comparaison des sections.
-
-
- Tableau I. L'évolution démographique de la population.
- Tableau II. Répartition ethnique de la population.
- Tableau III. Échantillon et population étudiés. Population can.-fr. (18 à 65 ans)
- Tableau IV. Groupes sanguins ABO du Saguenay-Lac Saint-Jean.
- Tableau V. Groupes sanguins A B O : comparaisons.
- Tableau VI. Fréquences moyennes des phénotypes A B O en France.
- Tableau VII. Groupes sanguins Rh (D) du Saguenag-Lac Saint-Jean.
- Tableau VIII. Groupes Rh standards.
- Tableau IX. Phénotypes A B O : répartition par sections et homogénéité des séries. (* P >0,10 ; ** P ≥0,10 ; *** P ≥0,50.)
- Tableau X. Groupes sanguins AB O : fréquences géniques par sections.
- Tableau XI. Répartition des valeurs de « p ».
- Tableau XII. Répartition des valeurs de « r ».
- Tableau XIII. Répartition des valeurs de « q ».
- Tableau XIV. Groupes sanguins Rh (D) : répartition par sections.
- Tableau XV. Répartition des valeurs Rh négatif.
- Tableau XVI. Groupes sanguins A B O : population britannique d'Arvida.
Introduction

Dans bien des domaines de l'anthropologie et de la génétique des populations, le Canada français offre à l'observateur un matériel exceptionnel (Laberge, 1966 ; Benoist et Auger, 1965). Peuplé depuis moins de trois siècles, ce très vaste territoire échappe en effet aux difficultés que suscite à toute recherche anthropologique le passé complexe et pratiquement indéchiffrable de bien des groupes humains. Les origines du peuplement, les courants migratoires, l'établissement des villes et des villages et la colonisation des zones vierges peuvent y être suivis à travers les documents historiques et les archives locales avec une précision bien difficile à égaler ailleurs. Aussi les hypothèses explicatives de l'anthropologue physique jouissent-elles dès le départ de conditions optimales : les causes de diversité et les facteurs extrinsèques de micro-évolution peuvent être saisis directement et non sous la forme approximative dont on est bien obligé de se contenter en général. Que l'on se place au niveau précis de petits groupes isolés, ou que l'on s'attache à une enquête sur l'ensemble de la population, les interférences avec des métissages inconnus, avec des origines incertaines ou avec une histoire régionale imprécise perturbent au minimum la présentation et l'interprétation des résultats. La connaissance très complète de l'histoire du peuplement et de l'histoire sociale nous invite à étudier la façon dont les mouvements humains qu'elles traduisent et les implantations dans les nouveaux milieux qu'elles impliquent se reflètent au sein du patrimoine génétique de populations humaines. Leur précision nous offre aussi une voie par laquelle on peut tenter d'aborder la question encore à peu près inconnue de la vitesse et du taux des changements de ce patrimoine.
Examiner les groupes sanguins d'une population implique une bonne définition préalable de cette population et de l'échantillon qui doit la représenter. Bien des travaux ont insisté sur le besoin de procéder pour cela d'une façon qui ne laisse pas les disparités régionales se fondre dans un ensemble moyen (Thieme, 1952 a ; 1952 b). Effectivement, en raffinant la délimitation des unités au sein desquelles est examinée la fréquence génique des groupes sanguins, on met en évidence des discontinuités dont certaines sont hautement significatives : les populations dont la fréquence moyenne a été déterminée par une enquête globale, même bien échantillonnées ne sont qu'imparfaitement connues tant qu'on n'a pas cherché si en leur sein n'existaient pas des sous-ensembles originaux qui traduisent une structuration, jusque-là méconnue, de cette population. Certes, l'unité théoriquement idéale serait la population panmictique quelles qu'en soient les dimensions, ou la petite population close aussi proche que possible du modèle de « l'isolat ». Mais la première est purement théorique, et justement les insuffisances de recherches trop globales tiennent au fait qu'elles impliquent à tort une telle homogénéité. À l'inverse, même dans les cas où existent des unités de dimension réduite qui se conforment assez étroitement aux caractéristiques de « l'isolat », on ne peut ni se contenter d'examiner seulement quelques-unes d'entre elles, ni se permettre de les étudier toutes d'une manière exhaustive. D'ailleurs, ce qu'apporte l'examen intensif de petites populations isolées à la connaissance des mécanismes évolutifs, pour précieux qu'il soit par sa précision, ne peut suffire et risque même de masquer des phénomènes dont la mise à jour requiert une perspective plus vaste. Les petites populations, en assurant la transmission du patrimoine génétique dans un ensemble où les individus, à travers toutes les circonstances de leur existence particulière, ont une influence très accrue par rapport à celle qu'ils exercent réellement dans les groupes plus vastes, voient leur évolution soumise à l'extrême aux fluctuations au hasard, et dissimulent ainsi l'amplitude et la direction des fluctuations orientées (Wright, 1967).
Aussi, le choix que nous avons fait de la dimension de notre étude tente-t-il de répondre aux quelques considérations présentées ci-dessus. Disposant de la possibilité d'étudier un territoire et un groupe ethnique bien défini (le Canada français et les Canadiens français), nous nous sommes d'abord préoccupés de reconnaître les quelques grands ensembles régionaux que l'histoire de leur peuplement permet de distinguer aisément. La colonisation des terres vierges où ont débarqué les premiers colons n'a en effet affecté qu'une part très faible de l'espace actuellement occupé par la population canadienne-française. Les premières colonies, établies le long du Saint-Laurent, essaimèrent vers les terres voisines. De ces terres partirent d'autres groupes de colons qui ouvrirent à l'exploitation des régions plus lointaines. On assiste ainsi à une série de mises en valeur régionales à partir de lieux d'émigration. C'est essentiellement sur cette base historique que nous avons appuyé notre subdivision en une série d'entités qui feront chacune l'objet d'un article.
Toutefois, au sein de ces régions, dont certaines sont très vastes, les colons n'ont pas non plus constitué des populations homogènes. Leur progression par étapes dans un milieu difficile a fragmenté les ensembles régionaux en sous-groupes plus ou moins isolés, au long des axes principaux de la migration.
Ces quelques considérations permettent de comprendre le choix que nous avons fait quant aux étapes de notre analyse. L'absence presque complète de données publiées sur les groupes sanguins des Canadiens français nous a conduits à présenter les résultats globaux des principales régions de peuplement. Mais pour éviter de laisser inaperçues des variations locales au sein de ces régions, nous avons poussé aussi avant que possible le morcellement géographique de nos résultats, en fonction de ce que nous apprenaient l'histoire régionale et les divisions administratives et sociales actuelles. Les résultats de cette fragmentation ne peuvent être qu'approximatifs : un certain arbitraire est en effet inhérent à tout essai de cet ordre. Partant du général pour aller vers les particularités, ce travail doit être complété d'une seconde étape qui prenne pour point de départ les cas particuliers qu'il met en évidence, de façon à pousser l'investigation de leurs caractéristiques propres et à tenter de les expliquer. Il doit donc être bien présent à l'esprit du lecteur que ce que nous présentons ici a pour but de voir s'il existe des disparités régionales et de localiser ces disparités.
Cette étude a été rendue possible grâce à la bienveillance de la Société canadienne de la Croix-Rouge de la ville de Québec et au concours de son directeur, le Colonel Louis A. Gagnon, qui nous ont permis (le recueillir les données indispensables à notre recherche, ainsi qu'à l'appui de l'Université de Montréal qui nous assura l'octroi nécessaire à la mise en marche de notre projet. Qu'ils en soient vivement remerciés.
I. LA RÉGION DU SAGUENAY-LAC SAINT-JEAN
ET SON PEUPLEMENT
Nous ne reprendrons pas ici les conditions générales du peuplement du Canada français, dont les conséquences génétiques et anthropologiques ont été très bien discutées par Laberge (1966), et nous concentrerons notre attention sur la région qui nous occupe ici : le Saguenay-Lac Saint-Jean (fig. 1).
Vaste territoire situé au Nord-Est de la partie habitée de la Province de Québec, cette région est nettement individualisée par ses caractères géographiques : elle appartient à une dépression du bouclier laurentien qui s'allonge du Sud-Est au Nord-Ouest, entre le fleuve Saint-Laurent qui lui est à peu près perpendiculaire et le bassin du Lac Saint-Jean. L'axe de cette dépression, la rivière Saguenay, permet aux eaux de ce très grand lac (44 km sur 34) de se déverser dans le Saint-Laurent. Les rives du lac et du Saguenay sont bordées de terres basses assez fertiles et l'ensemble de la région, dont la superficie est d'environ 15 330 km2, est cerné par les hauts plateaux de la forêt laurentienne. Les géographes (Blanchard, 1935) ont bien mis en évidence l'unité du Saguenay-Lac Saint-Jean et surtout son autonomie par rapport au reste de la Province.
La colonisation fut tardive. Le « royaume de Saguenay », dont l'existence avait été révélée à Jacques Cartier dès 1535 par des Indiens Montagnais (S.H.S. 1938), ne reçut d'autre implantation que celle d'un poste du Roi destiné à la traite des fourrures, situé à Chicoutimi. La Compagnie de la Baie d'Hudson interdit ensuite tout défrichement dans la région. Ce n'est qu'au printemps de 1838 qu'apparurent les premiers colons, des Canadiens français qui se concentrèrent principalement sur la rive sud du Saguenay et en particulier au fond de la Baie des Ha ! Ha ! À cette première vague de bûcherons qui progressèrent vers l'intérieur à mesure que les scieries s'y établirent, succéda une colonisation agricole plus systématique. De la même façon que dans les Laurentides ou dans certaines régions de l'Est du Québec, des curés jouèrent le rôle de pionniers en créant des établissements qui devinrent des paroisses : des sociétés de « défricheurs » se constituèrent et canalisèrent vers le Saguenay-Lac Saint-Jean l'important excédent démographique des paroisses anciennement établies (le la Province (surtout la région de Québec et les rives Nord et Sud de l'estuaire). Cette colonisation atteignit le Lac Saint-Jean en 1850, le contourna peu à peu, et vers 1885 l'occupation du sol était pratiquement accomplie (fig. 2).
Fig. 1.
Situation géographique du Saguenay-Lac Saint-Jean.
On assista à un essor démographique très rapide (tableau 1), fait pour une large part de la croissance naturelle des colons. On a noté un taux de naissance exceptionnel qui a pu monter en 1850 à 90%, (Blanchard, 1935). Quoique beaucoup plus basse de nos jours, la natalité y demeure encore l'une des plus fortes du Québec.
Fig. 2.
Les grandes étapes du peuplement

Tableau 1
L'évolution démographique de la population.
|
Années
|
|
|
|
|
|
|
1839 (1)
|
|
|
|
|
|
|
1851
|
|
|
|
|
|
|
1861 (2).
|
|
|
|
|
|
|
1871
|
|
|
|
|
|
|
1881
|
|
|
|
|
|
|
1891
|
|
|
|
|
|
|
1901
|
|
|
|
|
|
(1) Les chiffres de 1839 et 1851 proviennent de S.H.S., 1938.
(2) Les années 1861 à 1901 sont tirées des recensements décennaux du Canada.
(3) Les années 1911 à 1961 sont tirées des Annuaires Statistiques du Québec, 1956-1957 et 1964-1965.
|
Ainsi, l'essaimage de paroisses anciennes, surpeuplées par la suite de leur exceptionnelle croissance naturelle, vint-il se couler dans ce moule géographique encore vacant, et la population actuelle porte la trace à la fois de ces origines démographiques et des conditions que le cadre naturel imposa à son implantation.
Presque exclusivement canadienne-française (tableau II), cette population reproduit à une échelle réduite l'aventure des premiers colons français et de leurs descendants. Par cela même, elle voit l'intérêt de son étude se doubler de celui que lui confère cette situation exemplaire.
Tableau II
Répartition ethnique de la population.
|
Canadiens français
|
251 945
|
96,0%
|
|
D'origine étrangère
|
9 360
|
3,6%
|
|
Indiens
|
1 121
|
0,4%
|
II. LES GROUPES SANGUINS ABO ET RH
A. L'échantillon.
Nous avons exploité les données de la Croix-Rouge canadienne. Certes, immédiatement, une telle origine affecte nos résultats et notre échantillon de quelque suspicion, en raison des diverses critiques dont de telles enquêtes peuvent faire justement l'objet (non-représentativité, excès de groupes O, etc.). Toutefois, les causes qui rendent ces critiques méritées peuvent être évitées si l'on s'entoure de quelques précautions.
Les séries présentées ici proviennent des registres de la ville de Québec et portent les résultats des donneurs bénévoles qui ont participé aux « cliniques de sang » au cours de tournées de collecte effectuées par la Croix-Rouge de Québec entre 1962 et 1965. Voici donc les caractères de l'échantillon étudié :
1˚ Tous nos sujets en sont à leur premier don de sang, ce qui élimine l'une des causes d'erreur les plus fréquemment invoquées.
2˚ Nous avons relevé 10759 sujets. Par la suite ont été éliminés, en raison de leurs origines : 106 sujets examinés en dehors de la région étudiée, 408 étrangers, 472 religieux et religieuses dont l'origine ne pouvait être connue de nous. Ont également dus être comptés à part 30 sujets dont nous n'avons pu identifier le lieu de résidence et 54 sujets de la zone de Desbiens, qui ne pouvaient être rattachés à une autre zone. Les 9 689 Canadiens français restants forment un pourcentage appréciable de la population totale. Si, par ailleurs, nous tenons compte du fait qu'il s'agit de donneurs de sang, ce qui exclut les enfants et les vieillards, c'est en réalité un échantillon de la population adulte (tableau III) qui a été examiné. D'un point de vue strictement quantitatif, cet échantillon se place donc bien au-delà des exigences les plus rigoureuses. Il en va de même quant à sa composition régionale où il n'existe qu'une légère distorsion.
3˚ La composition qualitative de l'échantillon nous montre une sous-représentation du sexe féminin (environ 18%). Tout ce que nous savons sur les groupes sanguins indique qu'il ne s'agit pas là d'une influence véritablement perturbatrice. Effectivement, après avoir décompté séparément hommes et femmes, nous avons comparé la répartition des groupes ABO chez les 7 881 hommes et les 1808 femmes, sans mettre en relief de différence significative. L'âge est assez homogène en raison du recrutement. Il a été noté pour chaque sujet, mais nous ne tenterons pas ici d'en tenir compte, suivant en cela les travaux de la majorité (les auteurs.
Tableau III
Échantillon et population étudiés.
|
Sections
|
Population can.-fr.
(18 à 65 ans)
|
|
% de la population étudiée
|
|
|
|
|
|
|
|
|
1.
|
|
|
|
|
|
|
2.
|
|
|
|
|
|
|
3.
|
|
|
|
|
|
|
4.
|
|
|
|
|
|
|
5.
|
|
|
|
|
|
|
6.
|
|
|
|
|
|
|
7.
|
|
|
|
|
|
|
8.
|
|
|
|
|
|
|
9.
|
|
|
|
|
|
|
10.
|
|
|
|
|
|
|
11.
|
|
|
|
|
|
|
12.
|
|
|
|
|
|
|
13.
|
|
|
|
|
|
|
14.
|
|
|
|
|
|
|
15.
|
|
|
|
|
|
|
Ensemble
|
|
|
|
|
|
Le plus difficile tient à l'origine des sujets. Il s'agit, d'une part, de déceler ceux qui peuvent être comptés comme Canadiens français, et, d'autre part, de les situer géographiquement au sein de la région.
Ont été retenus comme Canadiens français les individus dont le nom ne laissait aucun doute quant à leur identité ethnique, et ceux qui portaient des patronymes étrangers introduits lors du mariage d'un ancêtre étranger et d'une femme canadienne-française voilà plus de quatre générations, c'est-à-dire avant 1850. Les descendants de ces unions se sont alors intégrés au groupe canadien-français. Nous avons pu découvrir dans le « Recueil de généalogie des comtes de Charlevoix et Saguenay », de G. Eloi (1941), la liste de quinze de ces patronymes, et en cas de doute nous avons préféré compter le sujet hors du groupe canadien-français, mais de tels cas sont rares. Ces aspects de l'analyse de l'échantillon soulignent bien l'avantage d'une population comme la nôtre et l'apport à la recherche que constitue la précision de son histoire.
La localisation (les sujets a demandé des choix plus délicats. Toute étude de la répartition géographique de groupes sanguins vise à déterminer la fréquence génique de ces caractères dans divers groupes locaux de la population qu'on peut considérer comme « autochtone ». On peut (Vallois et Marquer, 1964) se contenter du lieu de naissance pour déterminer l'implantation des sujets. Certains auteurs (Ranque et coll., 1958, par exemple) considèrent cela comme insuffisant et exigent le lieu de naissance des parents et des grands-parents. En fait, toute limite est conventionnelle dans les sociétés industrielles, et particulièrement dans les villes, lieu de régression démographique dont on sait que l'apport des campagnes vient en permanence modifier la composition. C'est la situation démographique locale, l'histoire du peuplement et les conditions socio-culturelles (migrations internes, choix du conjoint, changements économiques) qui peuvent seules nous indiquer le niveau auquel nous devons définir la localisation des sujets. Dans la région du Saguenay-Lac Saint-Jean nous avons ainsi été amenés à considérer que le lieu de résidence nous donnait une image étroitement fidèle des lieux de naissance et d'origine de nos sujets. Nous avons constitué autant de groupes locaux qu'il existait d'agglomérations résidentielles identifiées. Ce n'est que par la suite que nous avons rassemblé ces agglomérations en zones plus vastes, pour des raisons historiques ou méthodologiques qui seront commentées plus loin.
B. Les résultats.
1˚ Le système ABO.
Les auteurs traitant des origines du peuple français en Amérique sont d'accord pour affirmer que toutes les provinces françaises ont participé à l'hérédité des Canadiens français, mais on insiste pour souligner l'apport prépondérant de la Normandie (Langlois, 1935), de l'Île-de-France et du Centre-Ouest : Poitou, Aunis, Saintonge (Godbout, 1966).
TABLEAU IV
Groupes sanguins ABO du Saguenay-Lac Saint-Jean.
|
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Le tableau V présente les caractéristiques sérologiques de la France en général ainsi que celles des provinces mentionnées plus haut. Nous y joignons les fréquences des Indiens Montagnais-naskapi du Québec et celles des pays d'origine des Canadiens anglais. Nous avons recalculé, quand il y avait lieu, les fréquences p, q et r à partir des données phénotypiques, afin de faciliter la comparaison avec notre propre statistique. Le test d'homogénéité a été pratiqué sur les fréquences caractérisant les cinq provinces françaises regroupées ainsi que sur l'ensemble amérindien : pour les Français, le x2 = 0,563, soit une homogénéité avec P = 0,40, pour un degré de liberté ; la série amérindienne est homogène avec x2 = 0,965, soit P = 0,30.
Tableau V
Groupes sanguins A B O : comparaisons.
|
|
|
|
|
|
|
|
France (VALLOIS et MARQUER, 1964).
|
|
|
|
|
|
|
|
|
|
|
|
|
Provinces françaises regroupées (d'après VALLOIS, 1964).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Provinces françaises regroupées
|
|
|
|
|
Royaume-Uni (LABERGE, 1966)
|
|
|
|
|
Montagnais-naskapi (BLUMBERG et Coll., 1964)
|
|
|
|
|
Les chiffres du tableau V sont très démonstratifs : les fréquences caractérisant les deux séries françaises ainsi que les échantillons amérindien et britannique divergent grandement de celles des Canadiens français du Saguenay-Lac Saint-Jean.
La France se distingue nettement de la population canadienne-française avec un x2 très significatif de 89,985 ! (pour 3 ddl, au seuil de 0,01 : x2 = 11,345). Mais ce qui est encore plus intéressant, c'est la différence aussi nette que l'on trouve entre l'ensemble des provinces françaises ayant participé de façon prépondérante au peuplement de la Nouvelle-France et notre population québécoise d'aujourd'hui : le même test de comparaison donne un x2 = 30,358 ; quoique plus faible, la différence demeure significative avec une probabilité de plus de 99%.
La formule ABO des Canadiens français du Saguenay-Lac Saint-Jean tranche sur celle de l'ensemble de la population française par une élévation des fréquences du groupe A et surtout du groupe B, et par un abaissement marqué de la fréquence du groupe O. En effet, si nous considérons les limites établies par Vallois et Marquer (1964) (tableau VI) dans la majorité des départements français, nous constatons, d'une part, que la fréquence du phénotype B des Canadiens français (9,2%) se situe à la limite supérieure des valeurs françaises moyennes, et que la fréquence de O (39,6%) y est inférieure aux valeurs moyennes « à tendance faible » de Vallois et Marquer.
Fig. 3
Répartition de 5 populations suivant la méthode de Streng :
groupes sanguins ABO (explications dans le texte).
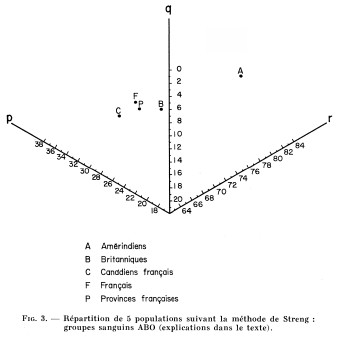
TABLEAU VI
Fréquences moyennes des phénotypes A B O en France.
Par conséquent, et tout en demeurant à l'intérieur des limites extrêmes des valeurs françaises régionales, la population française du Saguenay-Lac Saint-Jean fait nettement figure de population marginale. Par ailleurs, et contrairement à ce qu'on aurait pu prévoir, c'est la formule sanguine française qui se situe en position intermédiaire, entre celle des Indiens du Québec et celle des Canadiens français (voir fig. 3).
Une distance considérable sépare nos quatre populations d'origine européenne des Amérindiens. On doit souligner que notre population canadienne-française est à l'opposé de ce qu'un métissage avec les Indiens aurait pu produire, et que cette constatation est renforcée par le fait que le seul groupe que l'on puisse craindre de voir surestimer dans un échantillon de donneurs est le groupe O.
2˚ Le système Rh.
Les résultats ne portent que sur Rh (D).
Tableau VII
Groupes sanguins Rh (D) du Saguenag-Lac Saint-Jean.
Comparaisons.
Nous présentons au tableau VIII les chiffres de Moullec et Khérumian (Mourant, 1954) sur les fréquences phénotypiques du système Rh chez les étudiants parisiens ainsi, qu'à titre d'indication, les fréquences d'un groupe d'Algonquins du Québec (Chown et Lewis, 1956). Les fréquences phénotypiques déterminant les groupes Rh standards des Indiens Cree de la Baie James ont été calculées à partir des chiffres de Chown et Lewis (1956) sur les sous-groupes rhésus de ces Amérindiens.
Tableau VIII
Groupes Rh standards.
|
|
|
|
|
|
France (Région parisienne)
|
|
|
|
|
|
|
|
|
|
Indiens Cree (Baie James)
|
|
|
|
|
|
|
|
|
En France, d'après la statistique de Mourant (1954), le taux du phénotype Rh- pour la presque totalité du pays oscillerait entre 16% et 20%. La fréquence de notre série (Rh- = 15,8%) se situerait donc à la limite inférieure des valeurs françaises. Néanmoins, la comparaison entre les chiffres parisiens et ceux du Saguenay-Lac Saint-Jean indique une différence nulle
avec 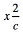 = 0,100 ( = 3,841 : avec P = 0,95 pour 1 ddl). = 0,100 ( = 3,841 : avec P = 0,95 pour 1 ddl).
Cette dernière constatation laisserait croire qu'au cours de l'histoire naturelle des Canadiens français, la distribution des facteurs Rh (D) aurait conservé une stabilité qu'on ne retrouve pas dans la distribution des groupes ABO.
III. LA RÉPARTITION RÉGIONALE
DES GROUPES SANGUINS AU SAGUENAY-LAC SAINT-JEAN
Après ces premières constatations, qui nous ont montré les particularités sérologiques de la population du Saguenay-Lac Saint-Jean en face de sa souche française, nous allons descendre à un niveau local, pour déceler l'existence éventuelle de variations géographiques au sein de cette population.
A. La division du territoire.
Les divisions administratives actuelles sont récentes et purement arbitraires : comtés électoraux découpés en 1931, municipalités qui ne représentent souvent qu'un fragment des unités que l'histoire et la géographie de la région nous invitent à considérer comme entité significative. Nous avons tenté de nous approcher au mieux d'un découpage qui corresponde aux grandes zones d'endogamie en suivant attentivement les conditions du premier peuplement et de l'essor qui l'a suivi.
La première forme de division administrative fut la « mission », territoire assez vaste, sillonné par un réseau de chemins et organisé autour des activités paroissiales qu'incarnait le curé. Pendant fort longtemps, ces « missions » gardèrent une large autonomie : jusqu'au début du XXe siècle, les communications étaient dangereuses, lentes, irrégulières. Trois saisons sur quatre, elles étaient le plus souvent impossibles. Les mariages contractés entre localités voisines, au sein d'une même « mission », créèrent ainsi des unités de population à partir de ces unités d'implantation et de peuplement.
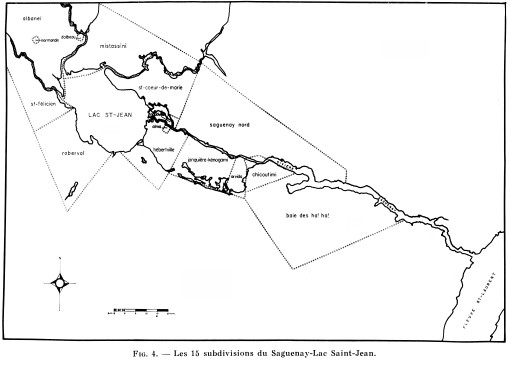
Fig. 4
Les 15 subdivisions du Saguenay-Lac Saint-Jean
Les travaux des ethnologues nous montrent que, longtemps après l'ouverture des réseaux de communication à large échelle, cette structure tend à persister, en particulier dans les régions peu propices aux migrations internes. Les villes voient, par contre, affluer des ruraux qui commencent à élargir le champ du choix du conjoint.
Tenant compte de ces données, nous avons été conduits à retenir pour notre étude 15 régions de base (fig. 4) dont la définition relève essentiellement des circonstances locales. Nous aurions aimé parfois pousser plus avant la subdivision, mais l'échantillon devenait alors trop exigu pour garder une valeur suffisante. Ces régions, qui couvrent la totalité du territoire du Saguenay-Lac Saint-Jean, regroupent chacune un nombre variable de municipalités, dont nous avons jugé utile de donner la liste en appendice.
1˚ La première « mission ».
La section 1 (Baie des Ha ! Ha !) correspond au premier noyau de colonisation établi dans la région. Au Sud de la rivière, elle représente le Bas et le Moyen Saguenay.
2˚ Zones urbaines.
Les sections 2, 3, 4, 6, 10 et 13 sont caractérisées par une faible surface et une haute densité démographique. Il s'agit de la série d'agglomérations nées du développement industriel. Point de convergence de la population rurale, chaque agglomération fut le lieu de fusion d'éléments de la région en général, et plus particulièrement des populations immédiatement avoisinantes. Aussi étions-nous tenus de faire de chacune de ces villes une section distincte.
À Chicoutimi, centre le plus ancien, se rattachent les localités de Rivière du Moulin et de Chicoutimi Nord, par la communauté de leur histoire (S.H.S., 1938) et leur proximité. Nous avons ajouté à cela la localité de Saint-Jean Eudes, après avoir constaté qu'elle ne différait pas de cette section du point de vue sérologique.
Les autres sections urbaines représentent soit des villes récentes (Arvida, Dolbeau, Alma) soit des centres administratifs et commerciaux (Jonquière-Kénogami, Roberval).
3˚ Zones rurales.
Quatre zones de terres fertiles, séparées par de mauvaises terres, constituent le véritable oekoumène agricole de la région du Saguenay-Lac Saint-Jean.
Dans notre division du territoire, ces quatre zones correspondent à autant de sections caractérisées par un complexe agricole ancien et stable (fig. 4) : Hébertville, plaine de Normandin ; petite plaine de Saint-Coeur-de-Marie et bande agricole de Saint-Félicien. Toutefois, nous avons dû subdiviser la plaine de Normandin en isolant le village de Normandin de l'ensemble des autres communautés de la plaine (groupées dans la section d'Albanel) en raison de l'hétérogénéité sérologique de ce village par rapport au reste de sa région.
4˚ Zones mixtes.
La section de Mistassini, immense terrasse sablonneuse entre les rivières Péribonka et Mistassini, regroupe des localités nées à la fin du siècle dernier du village de Mistassini et dévolues à une économie agricole et forestière.
La section 15 mériterait une étude plus détaillée, car elle regroupe les habitants d'une région accidentée qui, après un premier peuplement, a reçu une nouvelle immigration vers 1930. Cloisonnée par la géographie et diversifiée par l'histoire, cette zone devra faire l'objet d'études locales plus poussées.
La section 14 rassemble des centres résidentiels et industriels où la proportion d'étrangers est relativement forte.
B. Les groupes sanguins.
1˚ Système ABO.
La série de tableaux donnant la répartition des sections 1 à15 nous montre une diversité régionale impressionnante.
On constate que les valeurs du groupe O fluctuent entre un minimum de 36,2% (Albanel) et un maximum de 47,4% (Naudville). Si ces deux extrêmes correspondent à deux sections où notre échantillonnage apparaît légèrement faible (en nombre absolu), et en conséquence quelque peu suspect, les deux extrêmes de second rang [Baie des Ha ! Ha ! (36,6%) et Arvida (45,8%)] soulignent par l'importance de leur série l'existence réelle de cette variabilité des valeurs du groupe O. Dans l'ensemble des 15 sous-populations, 9 sections sont comprises entre 37% et 44%, alors que 3 sections ont moins de 37% et 3 dépassent 44%.
TABLEAU IX
Phénotypes A B O : répartition par sections et homogénéité des séries.
(* P >0,10 ; ** P ≥0,10 ; *** P ≥0,50.)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Section 1.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 2.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 3.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 4.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 5.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 6.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 7.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 8.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 9.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 10.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 11.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 12.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 13.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 14.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
|
Section 15.
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
Fréquences observées
|
|
|
|
|
|
|
Fréquences théoriques
|
|
|
|
|
|
|
Test d'homogénéité
|
|
Le groupe A se répartit entre un minimum de 39,4% (Arvida) et un maximum de 51% (Baie des Ha ! Ha !), deux sections dont le volume de l'échantillon ne laisse aucun doute quant au fondement réel de cet écart. Pour ce deuxième groupe sanguin, 4 sections ne dépassent pas 43,9% ; un ensemble de 10 sections se dissémine entre 44 et 50% et une section connaît une valeur supérieure à 50%.
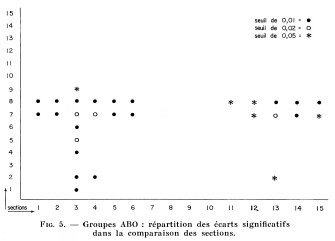
Fig. 5.
Groupes ABO : répartition des écarts significatifs
dans la comparaison des sections.
Pour le groupe B, la valeur minimale est de 6,3% (Naudville) et la valeur maximale de 17,2% (Normandin). Ces valeurs ne sauraient toutefois être acceptées que provisoirement, étant donné l'importance relativement faible de notre série de Naudville et l'hétérogénéité probable de notre série de Normandin. Deux sections ont des fréquences inférieures à 7% ; 10 sont comprises entre 7% et 11%, et 3 sections se situent au-dessus de 11%.
Le calcul du test d'homogénéité (x2 avec 1 ddl) montre que, pour une forte majorité des sous-populations du Saguenay-Lac Saint-Jean (14 sections sur 15), les distributions des valeurs observées et théoriques sont en accord nettement satisfaisant avec une probabilité toujours supérieure à 0,10. Le seul cas marginal est celui de Normandin (section 8) avec un x2 = 6,543 : une homogénéité à la limite P - 0,01. Toutefois, étant donné le fait que cette section 8 correspond à un unique petit village (notre série, quoique de 180 individus, représente 21,1% de la population étudiée) de type rural-agricole et où la population établie depuis plus de trois quarts de siècle est en écrasante majorité canadienne-française, nous sommes enclins à croire que le hasard d'échantillonnage serait en cause.
Le calcul des fréquences géniques (tableau X), par ailleurs, vient corroborer nos premières constatations quant aux tendances générales de la répartition des fréquences phénotypiques.
Tableau X
Groupes sanguins A B O : fréquences géniques par sections.
|
Sections
|
|
|
|
|
|
|
|
|
|
1.
|
|
|
|
|
|
|
|
|
|
2.
|
|
|
|
|
|
|
|
|
|
3.
|
|
|
|
|
|
|
|
|
|
4.
|
|
|
|
|
|
|
|
|
|
5.
|
|
|
|
|
|
|
|
|
|
6.
|
|
|
|
|
|
|
|
|
|
7.
|
|
|
|
|
|
|
|
|
|
8.
|
|
|
|
|
|
|
|
|
|
9.
|
|
|
|
|
|
|
|
|
|
10.
|
|
|
|
|
|
|
|
|
|
11.
|
|
|
|
|
|
|
|
|
|
12.
|
|
|
|
|
|
|
|
|
|
13.
|
|
|
|
|
|
|
|
|
|
14.
|
|
|
|
|
|
|
|
|
|
15.
|
|
|
|
|
|
|
|
|
La fréquence « p » se répartit entre une valeur minimale de 25,34% (Arvida) et une valeur maximale de 32,85% (Baie des Ha ! Ha !). Compte tenu de la fréquence du groupe A pour l'ensemble de notre échantillon canadien-français (30,15%), le classement (tableau XI) des 15 subdivisions qui lui sont sous-jacentes distingue quatre catégories de sous-populations.
Tableau XI
Répartition des valeurs de « p ».
|
Fréquences faibles (x à 27,99%)
|
4 sections.
|
|
Fréquences moyennes faibles (28 à 29,99%)
|
2 sections.
|
|
Fréquences moyennes fortes (30 à 31,99%)
|
8 sections.
|
|
Fréquences fortes (32% et plus)
|
1 section.
|
Cette distribution quelque peu hétérogène de nos sections semblerait indiquer qu'à une majorité de sous-populations semblables s'opposerait une catégorie de sous-populations se caractérisant par un commun abaissement de la fréquence du groupe A.
La fréquence « r », pour sa part, passe d'un minimum de 60,73% (Baie des Ha ! Ha !) à un maximum de 68,14% (Naudville). Le classement des 15 valeurs en quatre groupes répartis de part et d'autre de la fréquence du groupe O (62,86%) de l'ensemble des Canadiens français examinés est présenté au tableau XII.
Tableau XII
Répartition des valeurs de « r ».
|
Fréquences faibles (x à 60,99%)
|
1 section.
|
|
Fréquences moyennes faibles (61 à 62,99%)
|
8 sections.
|
|
Fréquences moyennes fortes (63 à 64,99%)
|
3 sections.
|
|
Fréquences fortes (65% et plus)
|
3 sections.
|
Enfin, les valeurs de « q » oscillent entre un minimum de 4,78% (Saint-Cœur-de-Marie) et un maximum de 10,79% (Saint-Félicien). Si on considère que la fréquence du groupe B de l'ensemble de nos sujets du Saguenay-Lac Saint-Jean est de 6,99%, le classement (tableau XIII) de notre série de sous-populations est le suivant :
Tableau XIII
Répartition des valeurs de « q ».
|
Fréquences faibles (x à 3,99%)
|
0 section.
|
|
Fréquences moyennes faibles (4 à 6,99%)
|
8 sections.
|
|
Fréquences moyennes fortes (7 à 9,99%)
|
5 sections.
|
|
Fréquences fortes (10% et plus)
|
2 sections.
|
2˚ Le système Rh.
Les résultats de notre statistique des groupes sanguins Rh standards des 15 sections du Saguenay-Lac Saint-Jean (tabl. XIV) semblent corroborer la principale constatation à laquelle nous avait conduit la répartition des groupes ABO.
Tableau XIV
Groupes sanguins Rh (D) : répartition par sections.
|
section
|
|
|
|
|
|
|
|
|
|
|
1.
|
|
|
|
|
|
|
2.
|
|
|
|
|
|
|
3.
|
|
|
|
|
|
|
4.
|
|
|
|
|
|
|
5.
|
|
|
|
|
|
|
6.
|
|
|
|
|
|
|
7.
|
|
|
|
|
|
|
8.
|
|
|
|
|
|
|
9.
|
|
|
|
|
|
|
10.
|
|
|
|
|
|
|
11.
|
|
|
|
|
|
|
12.
|
|
|
|
|
|
|
13.
|
|
|
|
|
|
|
14.
|
|
|
|
|
|
|
15.
|
|
|
|
|
|
La répartition de la fréquence phénotypique de Rh négatif passe d'un minimum de 12,7% (Baie des Ha ! Ha !) à un maximum de 18,4% (Roberval), deux cas dont la représentativité est étayée par un échantillon dont les nombres absolus et relatifs des sujets sont fort satisfaisants. En nous rappelant la fréquence (15,8%) du phénotype Rh négatif pour l'ensemble de notre série du Saguenay-Lac Saint-Jean, nous avons pu établir (tableau XV) un classement des valeurs phénotypiques de nos 15 sections
Tableau XV
Répartition des valeurs Rh négatif.
|
Fréquences faibles (x à 13,9%)
|
3 sections.
|
|
Fréquences moyennes faibles (14 à 15,9%%)
|
2 sections.
|
|
Fréquences moyennes fortes (16 à 17,9%)
|
5 sections.
|
|
Fréquences fortes (18% et plus)
|
5 sections.
|
3˚ Comparaisons entre les régions.
a) Groupes sanguins A B O. - Les sous-populations du Saguenay-Lac Saint-Jean ont été comparées deux à deux à l'aide du coefficient x2 de Pearson : un certain nombre de sections, pour 3 degrés de liberté, ont un écart significatif à un taux de confiance supérieur à 99%. Nous avons synthétisé les résultats en un graphique (fig. 5) destiné à mettre en relief les sections significativement différentes.
On remarque d'abord que les trois grandes agglomérations urbaines du Haut Saguenay, les sections 2, 3 et 4, sont très significativement différentes les unes des autres. Les fréquences « p » des sections de Chicoutimi et de Jonquière-Kénogami sont identiques et c'est principalement la haute fréquence de « q » de Jonquière-Kénogami qui les oppose, ce qui s'accorderait avec le fait que Chicoutimi correspond à un mélange plus général des éléments de la région.
Mais ce qui s'impose le plus à notre attention, c'est la présence de trois Sections, les sections 3, 7 et 8, qui s'écartent de façon significative de la plupart des autres.
La ville d'Arvida (section 3), située sur la rive sud du Haut Saguenay, se caractérise par la fréquence du groupe A, la plus faible de la région, et par des fréquences de B et de 0 très élevées. Toutefois, notre statistique (tableau XVI) d'une série parfaitement homogène (P = 100%) de 142 étrangers d'Arvida (britanniques d'après les patronymes) nous a révélé une formule sanguine dont l'abaissement manifeste de la valeur de « p » et l'augmentation très forte de « q » et surtout de « r »nous laisseraient le droit de penser que la formule sanguine singulière des Canadiens français d'Arvida pourrait tenir à un échange plus accentué avec la population étrangère (Arvida se compose d'une proportion relativement forte d'étrangers).
TABLEAU XVI
Groupes sanguins A B O : population britannique d'Arvida.
|
|
|
|
|
|
|
|
Nombres observés
|
|
|
|
|
|
|
Nombres théoriques
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Situées au Nord-Ouest du Lac Saint-Jean, les sections soeurs de Saint-Félicien et de Normandin (sections 7 et 8) sont également remarquables. Si la population de cette zone se distingue de celle de la plus grande partie du territoire, on remarque une absence de différence significative entre elle et la population de la zone qui lui est immédiatement voisine à l'Est (réf. à fig. 5).
Toutes deux membres d'une zone rurale agricole où la proportion canadienne-française de la population est nécessairement supérieure à celle d'Arvida, ces deux sections se caractérisent de façon générale par une fréquence moyenne de 0, une fréquence de A relativement faible et la fréquence du groupe B la plus forte de la région. En considérant que la section 7 possède une fréquence du groupe 0 relativement plus faible, il est intéressant de noter que, par leur très forte fréquence de B, nos deux sections conservent tout de même un commun dénominateur.
b) Le système Rh permet deux constatations : il existe là aussi des écarts significatifs entre section, mais ils sont bien moins nombreux ; ces écarts ne correspondent nullement aux sections qui ont été mises en relief par le système ABO.
Ces résultats ont certes un intérêt, car ils ne sont pas en contradiction avec des fluctuations indépendantes de deux systèmes géniques génétiquement indépendants. Nous devons toutefois honnêtement constater que, dans l'assertion de la signification de telles différences, nous parlons nécessairement en termes de probabilités. Même si aucun autre facteur que le hasard n'intervenait, un certain nombre de différences seraient significatives au niveau que nous demandons. Sur un nombre aussi considérable de comparaisons deux à deux, nous devons donc nous attendre à quelques différences dues au hasard et situées cependant individuellement à un niveau de confiance théoriquement acceptable. Cela abaisse donc beaucoup l'intérêt des constatations que nous faisons pour le système Rh qui, à la différence du système ABO, ne comprend pas un nombre d'écarts significatifs suffisants pour que nous puissions leur attacher une valeur ferme.
CONCLUSIONS
Ce travail répond à deux ordres de questions :
- apporter des données sur la population canadienne-française, jusqu'ici peu explorée quant à sa sérologie ;
- pousser au maximum la précision d'une enquête de répartition régionale des groupes sanguins en profitant des avantages considérables qui s'offrent dans cette population en raison de ses conditions d'implantation et de développement.
L'étendue du territoire où vivent les Canadiens français exige que cette recherche soit présentée en une série de monographies régionales, série que cet article inaugure. Dans la région qu'il décrit (Saguenay-Lac Saint-Jean), on constate :
- une différence nette avec les groupes sanguins généralement relevés en France, qui se traduit par une augmentation relativement importante du phénotype B ;
- une diversité intrarégionale assez profonde qui confirme le bien-fondé des recommandations de Thieme (1952) quant à l'importance de l'exacte représentativité géographique d'un échantillon dans la description de la population d'un territoire. Il semble que ce soit essentiellement une augmentation de B aux dépens de 0, significative dans trois sections sur 15, qui caractérise cette variation interne de la région du Saguenay-Lac Saint-Jean. Nous ne pouvons dépasser le niveau d'une simple constatation avant d'avoir accumulé les résultats. Notre prochaine étude, sur la Gaspésie, permettra peut-être de juger s'il s'agit d'un phénomène plus général ;
- les régions qui diffèrent par les fréquences ABO ne diffèrent pas nécessairement quant au Rh(D), et les variations intrarégionales des deux systèmes sont indépendantes.
APPENDICE
Démographie des sous-populations du Saguenay-Lac Saint-Jean.
1. Baie des Ha ! Ha ! 19 318 habitants.
Canton Saint-Jean 1 459 habitants.
Canton Otis 825 habitants.
Canton Ferland aucun chiffre disponible.
Grande Baie 941 habitants.
Port-Alfred 9 066 habitants.
Bagotville 5 629 habitants.
Paroisse Laterrière 1 398 habitants.
2. Chicoutimi 50 145 habitants.
Rivière du Moulin 4 386 habitants.
Chicoutimi 31 657 habitants.
Chicoutimi Nord 11 229 habitants.
Saint-Jean-Eudes 2 873 habitants.
3. Arvida 14 460 habitants.
4. Jonquière-Kénogami 41 686 habitants.
Shipshaw 1282 habitants.
Kénogami 11 816 habitants.
Jonquière 28 588 habitants.
Saint-Cyriac aucun chiffre disponible.
5. Hébertville 7 392 habitants.
Saint-Gédéon 890 habitants.
Saint-Bruno 1 158 habitants.
Hébertville Station 1257 habitants.
Notre-Dame d'Hébertville 1 604 habitants.
Lac-à-La-Croix 521 habitants.
Saint-Jérôme 1962 habitants.
6. Roberval 11 033 habitants.
Chambord 1 188 habitants.
Lac Bouchette 911 habitants.
Roberval 7 739 habitants.
Sainte-Hedwidge 1 195 habitants.
7. Saint-Félicien 8 068 habitants.
Saint-Prime 659 habitants.
Saint-Félicien 5 133 habitants.
Notre-Dame de la Doré 2 276 habitants.
8. Normandin 1838 habitants.
9. Albanel 6 218 habitants.
Saint-Méthode 1 372 habitants.
Albanel 917 habitants.
Saint-Edmond 886 habitants.
Didyme 1 310 habitants.
Girardville 1 733 habitants.
10. Dolbeau 6 052 habitants.
11. Mistassini 8 995 habitants.
Saint-Stanislas 867 habitants.
Saint-Eugène 1 067 habitants.
Sainte-Elisabeth aucun chiffre disponible.
Mistassini 3 461 habitants.
Sainte-Jeanne-d'Arc 909 habitants.
Saint-Augustin 818 habitants.
Péribonka 891 habitants.
Saint-Ludger-de-Milot 982 habitants.
12. Saint-Cœur-de-Marie 5250 habitants.
Saint-Coeur-de-Marie 1 302 habitants.
Saint-Henri-de-Taillon 907 habitants.
L'Ascension 1 742 habitants.
Sainte-Monique 1 299 habitants.
13. Alma 13 309 habitants.
14. Naudville-Isle Maligne 6 545 habitants.
Naudville 4 475 habitants.
Isle Maligne 2 070 habitants.
15. Saguenay Nord 12 007 habitants.
Notre-Dame-du-Rosaire 615 habitants.
Saint-Léon 1 182 habitants.
Saint-Nazaire 1 978 habitants.
Canton Bourget 961 habitants.
Saint-Ambroise 1 576 habitants.
Canton Bégin 1 055 habitants.
Saint-David de Falardeau 1 312 habitants.
Saint-Honoré 1 009 habitants.
Saint-Jean-Vianney 640 habitants.
Saint-Fulgence 1 094 habitants.
Sainte-Rose du Nord 585 habitants.
BIBLIOGRAPHIE
ANNUAIRES STATISTIQUES. Province du Québec, 1956-1957, 1962, et 1964-1965. Bur. de la Stat. du Québec. Imprim. de la Reine, Éditeur, Québec.
BENOIST (J.) et AUGER (F.), 1965. Note sur l'anthropométrie des Canadiens français. Anthropologica, N.S., t. 7, pp. 5-11.
BLANCHARD (R.), 1935. Province de Québec : l'Est du Canada français. Ed. Beauchemin, Montréal, 328 p.
BLUMBERG (B. S.) et collaborateurs, 1964. Blood groups of the Naskapi and Montagnais Indians of Schefferville, Québec. Hum. Biol., t. 36, pp. 246-272.
CHOWN (B.) et LEWIS (M.), 1956. The blood group genes of the Cree Indians and the Eskimos of the Ungava district of Canada. Am. J. Phys. Anth., t. 14, pp. 215-224.
ELOI (G.), 1941. Recueil de généalogie des comtés de Charlevoix et Saguenay. Ed. Soc. Hist. Saguenay, Malbaie, 594 p.
GODBOUT (A.), 1946. Nos hérédités provinciales françaises. Archives de Folklore, vol. 1, Université Laval, Ed., Fides, Montréal.
LABERGE (C.), 1966. Prospectus for genetic studies in the French Canadians, with preliminary data on blood groups and consanguinity, Bull. of John Hopkins Hosp., t. 118, pp. 52-68.
LANGLOIS (G.), 1935. Histoire de la population canadienne-française. Ed. Albert Lévesque, Montréal, 309 p.
MAGNAN (C.), 1967. Répartition des groupes sanguins ABO et Rh (D) au Saguenay-Lac Saint-Jean. Thèse M.A. dactylographiée, département d'anthropologie de l'Université de Montréal, 96 p., 6 fig., 18 tabl., 4 append.
MOURANT (A.E.), 1954. The distribution of the human blood groups. Charles C. Thomas Ed., Springfield, Ill, 438 p.
BANQUE (B.) et collaborateurs, 1958. Quelques réflexions sur la méthode séro-anthropologique en Europe occidentale à propos d'une enquête sur la population insulaire corse. Transfusion, t. 1, pp. 329-350.
RECENSEMENT DES CANADAS, 1863. Années 1860-1861. Québec, S. B. Foote, Ed.
RECENSEMENT DU CANADA, 1873. Années 1870-1871. Ottawa, 1. B. Taylor, Ed.
Id., 1882. Années 1880-1881. Ottawa, R. Maclean, Ed.
Id., 1893. Années 1890-1891. Ottawa, Ed. Impr. du Roi.
Id., 1903. Année 1901. Ottawa, Ed. Impr. du Roi.
S.H.S., 1938. L'Histoire du Saguenay. Depuis l'origine jusqu'à 1870. Ed. Soc. Hist. Saguenay, Chicoutimi, 331 p.
THIEME (F.), 1952 a. The geographic and racial distribution of ABO and Rh blood types and tasters of PTC in Puerto Rico. Am. J. Hum. Genet., t. 4, pp. 94-112.
Id., 1952 b. The population as a unit of study. Am. Anth., t. 54, pp. 504-509.
VALLOIS (H.V.) et MARQUER (P.), 1964. La répartition en France des groupes sanguins ABO. Bull. et Mém. Soc. d'Anthr. de Paris, t. 6, XIe s., pp. 1-200.
WRIGHT (S.), 1967. The foundations of population genetics in Heritage from Mendel. B. Alexander Brink Ed., Univ. of Wisconsin Press, pp. 245-263.
|